Пластичные взрывчатые вещества
Содержание:
- Функции пластид высших растений и их разнообразие[править | править код]
- См.Также
- Общие черты строения пластид высших растений
- Использование
- II.2 Характеристика бальзамов, эфирных масел, смол, камеди
- III. ПОЧКИ, ТИПЫ ПОЧЕК ПО ПРОИСХОЖДЕНИЮ. ПРИВЕДИТЕ РИСУНОК ВЕГЕТАТИВНОЙ ПОЧКИ В ПРОДОЛЬНОМ РАЗРЕЗЕ IV. В КРУПНОМ МАСШТАБЕ НАРИСУЙТЕ СЕМЯЗАЧАТОК С ЗАРОДЫШЕВЫМ МЕШКОМ В ПРОДОЛЬНОМ РАЗРЕЗЕ. ИЗ КАКИХ ЧАСТЕЙ СЕМЯЗАЧАТКА РАЗВИВАЮТСЯ ТЕ ИЛИ ИНЫЕ ЧАСТИ СЕМЕНИ
- V. СПОСОБЫ РАСПРОСТРАНЕНИЯ ПЛОДОВ И СЕМЯН. ПРИВЕДИТЕ ПРИМЕРЫ. БИОЛОГОИЧЕСКАЯ РОЛЬ РАСПРОСТРАНЕНИЯ ПЛОДОВ И СЕМЯН
- VI. ПОНЯТИЕ О ВИДЕ РАСТЕНИЙ. СОСТАВЛЕНИЕ ВИДОВЫХ НАЗВАНИЙ РАСТЕНИЙ СОГЛАСНО БИНАРНОМУ МЕТОДУ К. ЛИННЕЯ. ВЫПИШЕТЕ ИЗ «СПИСКА ОСНОВНЫХ СЕМЕЙСТВ И ВИДОВ» 6 ВИДОВ (ИЗ НИХ 2 ОДНОГО РОДА) ИЗ РАЗНЫХ СЕМЕЙСТВ
- VII. ЖИЗНЕННЫЙ ЦИКЛ ПЛАУНА БУЛАВОВИДНОГО И СЕЛАГИНЕЛЛЫ НАРИСУЙТЕ КОЛОСКИ И ЗАРОСТКИ ЭТИХ РАСТЕНИЙ
- VIII. ХАРАКТЕРИСТИКА СЕМ. БОБОВЫЕ (МОТЫЛЬКОВЫЕ). УКАЖИТЕ ВАЖНЕЙШИЕ ДИКОРАСТУЩИЕ И КУЛЬТУРНЫЕ РАСТЕНИЯ ИЗ ЭТОГО СЕМЕЙСТВА (НЕ МЕНЕЕ 15 ВИДОВ)
- IX. ПОНЯТИЕ О ЗОНАЛЬНОЙ И ИНТРАЗОНАЛЬНОЙ РАСТИТЕЛЬНОСТИ. ПРИВЕДИТЕ ПРИМЕРЫ
- Судьба варягов
- I.2 Фотосинтез, необходимые для него условия
- В каком оружии используется универсальный калибр?
- Примечания
- Геном и белоксинтезирующая система пластид высших растений[править | править код]
Функции пластид высших растений и их разнообразие[править | править код]
Пластиды высших растений способны к дифференцировке, дедифференцировке и редифференцировке, набор пластид в клетке зависит от её типа. Пластиды высших растений разнообразны по строению и выполняют широкий спектр функций:
- фотосинтез;
- восстановление неорганических ионов (нитрита, сульфата);
- синтез многих ключевых метаболитов (порфирины, пурины, пиримидины, многие аминокислоты, жирные кислоты, изопреноиды, фенольные соединения и др.), при этом некоторые синтетические пути дублируют уже существующие пути цитозоля;
- синтез регуляторных молекул (гиббереллины, цитокинины, АБК и др.);
- запасание железа, липидов, крахмала.
По окраске и выполняемой функции выделяют следующие типы пластид:
Растительные клетки листостебельного мха Plagiomnium affine с видимыми хлоропластами (сильно увеличено)
- Пропластиды — предшественники остальных типов пластид, присутствуют в меристематических клетках. Пропластиды имеют размеры от 0,2 до 1 мкм, что значительно меньше, чем размеры дифференцированных пластид. Внутренняя мембранная система развита слабо, содержат меньше рибосом чем дифференцированные пластиды, могут содержать отложения белка фитоферритина, основная функция которого хранение ионов железа.
-
Лейкопласты — неокрашенные пластиды, участвующие в синтезе изопреноидов эфирных масел (как правило моно- и сесквитерпенов). Характерной особенностью лейкопластов является наличие ретикулярного футляра — сети мембран гладкого эндоплазматического ретикулума, окружающей пластиду. Иногда под термином «лейкопласты» понимают любые неокрашенные пластиды, при этом выделяют следующие типы: амилопласты, элайопласты, протеинопласты.
- Амилопласты — внешне похожи на пропластиды, но в строме содержатся гранулы крахмала. Амилопласты, как правило, присутствуют в запасающих органах растений, в частности в клубнях картофеля. В грависенсорных клетках корня амилопласты играют роль статолитов. Амилопласты высших растений могут превращаться в хлоропласты или хромопласты.
- Элайопласты — служат для запасания жиров.
- Протеинопласты — служат для запасания белков.
- Этиопласты, или темновые пластиды, развиваются из пропластид в темноте, при освещении они превращаются в хлоропласты. В этиопластах отсутствует хлорофилл, но содержится большое количество протохлорофиллида. Липиды внутренних мембран стромы хранятся в форме рельефной мембранной структуры, называемой проламеллярным телом. Формирование квазикристаллической структуры проламеллярного тела происходит из-за отсутствия мембранных белков тилакоидов, необходимых для их формирования. Известно, что свет инициирует синтез белков тилакоидных мембран и хлорофилла из накопленного протохлорофиллида.
- Хлоропласты — зелёные пластиды, основной функцией которых является фотосинтез. Хлоропласты как правило имеют элипсовидную форму и длину от 5 до 8 мкм. Количество хлоропластов в клетке различно: в клетке хлоренхимы листа Arabidopsis содержится около 120 хлоропластов, в губчатой хлоренхиме листа клещевины их около 20, клетка нитчатой морской водоросли Spirogyra содержит единственный лентовидный хлоропласт. Хлоропласты имеют хорошо развитую эндомембранную систему, в которой выделяют тилакоиды стромы и стопки тилакоидов — граны. Зелёная окраска хлоропластов обусловлена высоким содержанием основного пигмента фотосинтеза — хлорофилла. Помимо хлорофилла хлоропласты содержат различные каротиноиды. Набор пигментов, участвующих в фотосинтезе (и, соответственно окраска) различен у представителей разных таксонов.
- Хромопласты — пластиды, окрашенные в жёлтый, красный или оранжевый цвет. Хромопласты могут развиваться из пропластид или повторно дифференцироваться из хлоропластов; также хромопласты могут редифференцироваться в хлоропласты. Окраска хромопластов связана с накоплением в них каротиноидов. Хромопласты определяют окраску осенних листьев, лепестков некоторых цветов (лютики, бархатцы), корнеплодов (морковь), созревших плодов (томат).
См.Также
Общие черты строения пластид высших растений
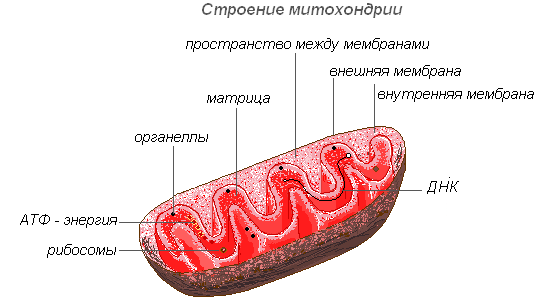
Типичные пластиды высших растений окружены оболочкой из двух мембран — внешней и внутренней. Внутренняя и внешняя мембраны пластид бедны фосфолипидами и обогащены галактолипидами. Внешняя мембрана не имеет складок, никогда не сливается с внутренней мембраной и содержит поровый белок, обеспечивающий свободный транспорт воды, ионов и метаболитов с массой до 10 кДа. Внешняя мембрана имеет зоны тесного контакта с внутренней мембраной; предполагается, что в этих участках осуществляется транспорт белков из цитоплазмы в начале пластид. Внутренняя мембрана проницаема для небольших незаряженных молекул и для недиссоциированных низкомолекулярных монокарбоновых кислот, для более крупных и заряженных метаболитов в мембране локализованы белковые переносчики. Строма — внутреннее содержимое пластид — представляет собой гидрофильный матрикс, содержащий неорганические ионы, водорастворимые органические метаболиты, геном пластид (несколько копий кольцевой ДНК), рибосомы прокариотического типа, ферменты матричного синтеза и другие ферментативные системы. Эндомембранная система пластид развивается в результате отшнуровки везикул от внутренней мембраны и их упорядочивания. Степень развития эндомембранной системы зависит от типа пластид. Наибольшего развития эндомембранная система достигает в хлоропластах, где она является местом протекания световых реакций фотосинтеза и представлена свободными тилакоидами стромы и тилакоидами, собранными в стопки — граны. Внутреннее пространство эндомембран называется люмен. Люмен тилакоидов, также как и строма, содержит ряд водорастворимых белков.
Использование
Дело в том, что бризантность (дробящий эффект) небольших зарядов взрывчатки быстро уменьшается при удалении от точки детонации. Грубо говоря, если десять грамм ВВ взорвется у вас в сжатом кулаке, то вы гарантировано лишитесь пальцев. Если же аналогичное количество взрывчатки сдетонирует в двадцати сантиметрах от вашей руки, то ущерб будет минимален. Вывод из этого прост: для нанесения максимального ущерба объекту взрывчатка должна находиться максимально близко к нему.
В этом отношении ПВВ идеален, заряд пластичной взрывчатки можно разместить не просто близко к разрушаемому объекту, а прилепить к нему. Металлическую балку или швеллер можно облепить ПВВ со всех сторон и этому не помешают выступы, болты или заклепки.
Да и крепить пластичную взрывчатку куда проще и быстрее, чем, например, тротиловые шашки.
II.2 Характеристика бальзамов, эфирных масел, смол, камеди
III.
ПОЧКИ, ТИПЫ ПОЧЕК ПО ПРОИСХОЖДЕНИЮ. ПРИВЕДИТЕ РИСУНОК ВЕГЕТАТИВНОЙ ПОЧКИ В
ПРОДОЛЬНОМ РАЗРЕЗЕ IV. В КРУПНОМ МАСШТАБЕ НАРИСУЙТЕ СЕМЯЗАЧАТОК С
ЗАРОДЫШЕВЫМ МЕШКОМ В ПРОДОЛЬНОМ РАЗРЕЗЕ. ИЗ КАКИХ ЧАСТЕЙ СЕМЯЗАЧАТКА
РАЗВИВАЮТСЯ ТЕ ИЛИ ИНЫЕ ЧАСТИ СЕМЕНИ
V.
СПОСОБЫ РАСПРОСТРАНЕНИЯ ПЛОДОВ И СЕМЯН. ПРИВЕДИТЕ ПРИМЕРЫ. БИОЛОГОИЧЕСКАЯ РОЛЬ
РАСПРОСТРАНЕНИЯ ПЛОДОВ И СЕМЯН
VI.
ПОНЯТИЕ О ВИДЕ РАСТЕНИЙ. СОСТАВЛЕНИЕ ВИДОВЫХ НАЗВАНИЙ РАСТЕНИЙ СОГЛАСНО
БИНАРНОМУ МЕТОДУ К. ЛИННЕЯ. ВЫПИШЕТЕ ИЗ «СПИСКА ОСНОВНЫХ СЕМЕЙСТВ И ВИДОВ» 6
ВИДОВ (ИЗ НИХ 2 ОДНОГО РОДА) ИЗ РАЗНЫХ СЕМЕЙСТВ
VII.
ЖИЗНЕННЫЙ ЦИКЛ ПЛАУНА БУЛАВОВИДНОГО И СЕЛАГИНЕЛЛЫ НАРИСУЙТЕ КОЛОСКИ И ЗАРОСТКИ
ЭТИХ РАСТЕНИЙ
VIII.
ХАРАКТЕРИСТИКА СЕМ. БОБОВЫЕ (МОТЫЛЬКОВЫЕ). УКАЖИТЕ ВАЖНЕЙШИЕ ДИКОРАСТУЩИЕ И
КУЛЬТУРНЫЕ РАСТЕНИЯ ИЗ ЭТОГО СЕМЕЙСТВА (НЕ МЕНЕЕ 15 ВИДОВ)
IX. ПОНЯТИЕ О ЗОНАЛЬНОЙ И ИНТРАЗОНАЛЬНОЙ
РАСТИТЕЛЬНОСТИ. ПРИВЕДИТЕ ПРИМЕРЫ
Судьба варягов
Государство Рюриковичей к X-XII стало очень богатым, и могло позволить себе просто «купить» нужных для службы варягов. Их оставляли в городских гарнизонах и дружине. Нападения викингов на русские города были бы делом бессмысленным. Легче было получить хорошее жалование за службу.
В городах простой народ с варягами часто не ладил – случались стычки. Вскоре ситуация начала выходить из-под контроля и Ярославу Владимировичу пришлось ввести «понятия» — Русскую правду. Так появился первый в истории России правовой документ.
В XII веке завершается эпоха викингов. На Руси упоминания о варягах исчезают из летописей уже к XIII веку, а русы растворяются в славянском русском народе.
I.2 Фотосинтез, необходимые для него условия
Фотосинтез у зеленых растений – это
процесс преобразования света в химическую энергию органических соединений,
синтезируемых из диоксида углерода и воды. Процесс фотосинтеза представляет
собой цепь окислительно-восстановительных реакций, совокупность которых
подразделяют на две фазы – световую и темновую.
1.
Световая
фаза. Для
этой фазы характерно то, что энергия солнечной радиации, поглощенная пигментами
системы хлоропластов, преобразуется в электрохимическую.
При действии света на хлоропласт
начинается электронный поток по системе переносчиков – сложных органических
соединений, встроенных в мембраны тилакоидов. С переносом электронов по ЭТЦ
сопряжено активное поступление протонов через тилакоидную мембрану из стромы
внутрь тилакоида. В тилакоидном пространстве происходит увеличение концентрации
протонов за счет расщепления молекул воды и в результате окисления электронного
переносчика пластохинона на внутренней стороне мембраны. Когда протоны идут
обратно по градиенту из тилакоидного пространства в строму, на наружной
поверхности тилакоида с участием фермента АТФ-синтетазы из АДФ и фосфорной
кислоты синтезируется АТФ, т. е. происходит фотосинтетическое фосфореилирование
с запасанием энергии в АТФ, которая затем переходит в строму хлоропласта.
Заканчивается передача электронов
следующим образом. Достигнув внешней поверхности мембраны тилакоида, пара
электронов следует с ионом водорода, находящимся в строме. Оба электрона и ион
водорода присоеденяются к молекуле переносчика водорода – НАДФ+
(никатиномидадениндинуклетидфосфат), который при этом переходит в свою
востановленную форму
НАДФ•Н+Н+:
НАДФ++2Н++2е-→НАДФ•Н+Н+
Следовательно активированные световой
энергией электроны используются на присоедининие атома водорода к его
переносчику, т. е. на восстановление НАДФ+ в НАДФ•Н+Н+,
который с наружной поверхности фотосинтетической мембраны переходит в
строму.
В молекулах хлорофилла,
утративших свои электроны, образовавшиеся электронные «дырки» действуют как
сильный окислитель и отрывают электроны от молекул воды. Через ряд переносчиков
эти электроны передаются на молекулу хлорофилла и заполняют «дырку». Внутри
тилакоида происходит фотоокислние (фотолиз) воды, в результате которого
выделяется свободный кислород, а также накапливаются ионы водорода
2Н2О→4Н++4е-+О2
Таким образом, во время световой фазы
фотосинтеза происходят три процесса: образование кислорода вследствие
разложения воды, синтез АТФ и образование атомов водорода в форме НАДФ•Н2.
Кислород диффундирует в атмосферу, а АТФ и НАДФ•Н2 транспортируются
в матрикс пластид и участвуют в процессе темновой фазы.
2.Темновая фаза фотосинтеза протекает в
матриксе хлоропласта как на свету, так и в темноте и представляет собой ряд
последовательных преобразований СО2, поступаещего из воздуха.
Осуществляются реакции темновой фазы за счет энергии АТФ и НАДФ•Н2 и
использовании имеющихся в пластидах пятиуглеродных сахаров, один из которых –
рибулозодифосфат – является акцептором СО2. Ферменты связывают
пятиуглеродный сахар с углекислым газом воздуха. При этом образуются соединения
которые последовательно восстанавливаются до шестиуглеродной молекулы глюкозы.
Суммарная реакция фотосинтеза
6СО2+6Н2 энергия света С6Н12О6+6О2
хлорофилл
В процессе фотосинтеза кроме
моносахаридов (глюкоза и др.), которые превращаются в крахмал и запасаются
растением, синтезируются мономеры других органических соединений – аминокислоты,
глицерин и жирные кислоты. Таким образом, благодоря фотосинтезу растительные, а
точнее – хлорофиллсодержащие, клетки обеспечивают себя и все живое на Земле
необходимыми органическими веществами и кислородом.
В каком оружии используется универсальный калибр?
Примечания
- >
Геном и белоксинтезирующая система пластид высших растений[править | править код]
Одним из доказательств происхождения пластид от древних цианобактерий служит схожесть их геномов, хотя пластидный геном (пластом) значительно меньше. Пластом высших растений представлен многокопийной кольцевой двуцепочечной ДНК (плДНК) размером от 75 до 290 тыс. п. н. В большинстве пластидных геномов присутствуют два инвертированных повтора (IRA и IRB), разделяющих молекулу ДНК на две уникальные области: большую (LSR) и малую (SSR). В инвертированных повторах содержатся гены всех четырёх рРНК (4,5S, 5S, 16S и 23S), входящих в состав пластидных рибосом, а также гены некоторых тРНК. Голосеменные и растения семейства Бобовые не содержат инвертированных повторов. Многие пластидные гены организованы в опероны — группы генов, считывающихся с общего промотора. Некоторые пластидные гены имеют экзон-интронную структуру. В пластидах кодируются гены, обслуживающие процессы транскрипции и трансляции (гены «домашнего хозяйства»), а также некоторые гены, обеспечивающие выполнение функций пластид в клетке, прежде всего фотосинтез.
Транскрипцию в пластидах обеспечивают РНК-полимеразы двух типов:
- Мультисубъединичная пластидная РНК-полимераза бактериального типа состоит из двух α-субъединиц и по одной β, β’, β» (все эти субъединицы кодируются в пластидном геноме). Однако для её активации необходимо присутствие σ-субъединицы, которая кодируется в ядре растительной клетки и импортируется в пластиды при освещении. Таким образом пластидная РНК-полимераза активна только на свету. Пластидная РНК-полимераза может обеспечивать транскрипцию с генов с эубактериальными промоторами (большинство генов фотосинтетических белков), а также с генов, имеющих универсальные промоторы.
- Мономерная РНК-полимераза фагового типа кодируется в ядре и белок имеет специальную сигнальную последовательность, обеспечивающую импорт в пластиды. Обеспечивает транскрипцию генов «домашнего хозяйства» (в частности гены rif-оперона, который содержит гены пластидной РНК-полимеразы).
Процесс созревания транскриптов пластид имеет свои особенности. В частности, пластидные интроны способны к автосплайсингу, то есть вырезание интронов происходит автокаталитически. Кроме того, в пластидах происходит редактирование РНК — химическая модификация оснований РНК, приводящая к изменению закодированной информации (наиболее часто происходит замена цитидина на уридин). Большинство зрелых мРНК пластид содержат в 3′-некодирующей области шпильку, защищающую её от рибонуклеаз.
Пластиды имеют рибосомы прокариотического типа с коэффициентом седиментации 70S (с меньшим количеством белков, по сравнению с эукариотическими рибосомами). Рибосомы содержат четыре типа рРНК, три из которых гомологичны эубактериальным 5S, 16S и 23S, а 4,5S рРНК гомологична 3′-участку 23S-рРНК.